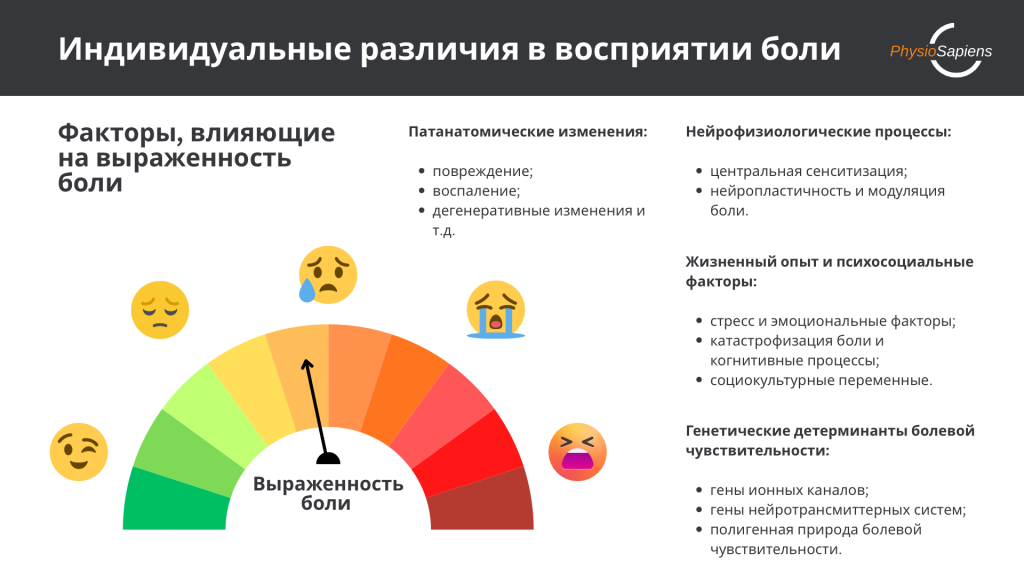
Восприятие боли представляет собой сложный многомерный феномен, характеризующийся выраженной межиндивидуальной вариабельностью. Клиническая практика регулярно демонстрирует парадоксальные ситуации: у одних пациентов значительные структурные повреждения, выявляемые при магнитно-резонансной томографии (МРТ), сопровождаются минимальной болью, в то время как у других интенсивная боль возникает при отсутствии явных патанатомических изменений [1], [2].
Современное понимание боли основывается на биопсихосоциальной модели, которая постулирует, что болевое восприятие формируется через взаимодействие биологических, психологических и социокультурных факторов [1], [2]. В рамках этой модели особое значение приобретает разделение восприятия боли на два ключевых домена: патанатомический и нейрофизиологический. Патанатомический домен включает структурные изменения тканей — воспаление, повреждение, дегенеративные процессы и т.д., которые могут быть подтверждены посредством методов инструментальной диагностики. Нейрофизиологический домен охватывает процессы центральной обработки ноцицептивной информации, включая центральную сенситизацию, нейропластичность, модуляцию боли в центральной нервной системе (ЦНС) и активацию специфических мозговых структур [11].
Накопленные данные указывают на то, что генетические факторы объясняют значительную долю межиндивидуальной вариабельности болевой чувствительности. Исследования близнецов и семейные исследования демонстрируют наследуемость болевых фенотипов, а молекулярно-генетические исследования идентифицировали множество генов-кандидатов, влияющих на ноцицепцию [3], [15], [24].
Параллельно с генетическими факторами на формирование болевого восприятия существенное влияние оказывает жизненный опыт индивида, включая психосоциальные переменные, стресс, эмоциональное состояние и социокультурный контекст [2], [9].
Цель данной статьи — представить интегративный анализ механизмов, лежащих в основе индивидуальных различий в восприятии боли, с акцентом на взаимодействие патанатомического и нейрофизиологического доменов, генетических детерминант и влияния жизненного опыта. Также мы разберем примеры несоответствия между данными МРТ и субъективным болевым опытом, что имеет критическое значение для понимания природы хронической боли и разработки персонализированных программ лечения.
Патанатомический домен боли

Патанатомический домен боли традиционно рассматривается как совокупность структурных изменений в тканях, которые могут служить источником ноцицептивной стимуляции. Эти изменения включают механические повреждения, воспалительные и дегенеративные процессы, опухолевые и другие морфологические аномалии, визуализируемые с помощью МРТ, компьютерной томографии (КТ) и других методов инструментальной диагностики.
Однако клинические наблюдения и исследования последних десятилетий демонстрируют слабую корреляцию между степенью патанатомических изменений и интенсивностью болевого синдрома. Структурные аномалии, такие как грыжи межпозвонковых дисков, дегенеративные изменения суставов или повреждения мягких тканей, могут обнаруживаться у асимптоматичных индивидов, в то время как пациенты с выраженной болью могут демонстрировать минимальные или отсутствующие патанатомические изменения [12].
Это несоответствие между патанатомией и болевым опытом указывает на то, что структурные изменения тканей являются лишь одним из компонентов сложной системы, определяющей болевое восприятие. Патанатомические изменения могут инициировать ноцицептивную сигнализацию, однако интенсивность и качество болевого опыта в значительной степени определяются процессами центральной обработки этой информации, что относится к нейрофизиологическому домену [11], [12].
Важно отметить, что патанатомический домен не является статичным. Хроническая боль может приводить к вторичным структурным изменениям в центральной нервной системе, включая изменения объема серого вещества [18], [20]. Эти изменения отражают нейропластические процессы, связанные с длительной ноцицептивной стимуляцией и центральной сенситизацией [28].
Нейрофизиологический домен боли
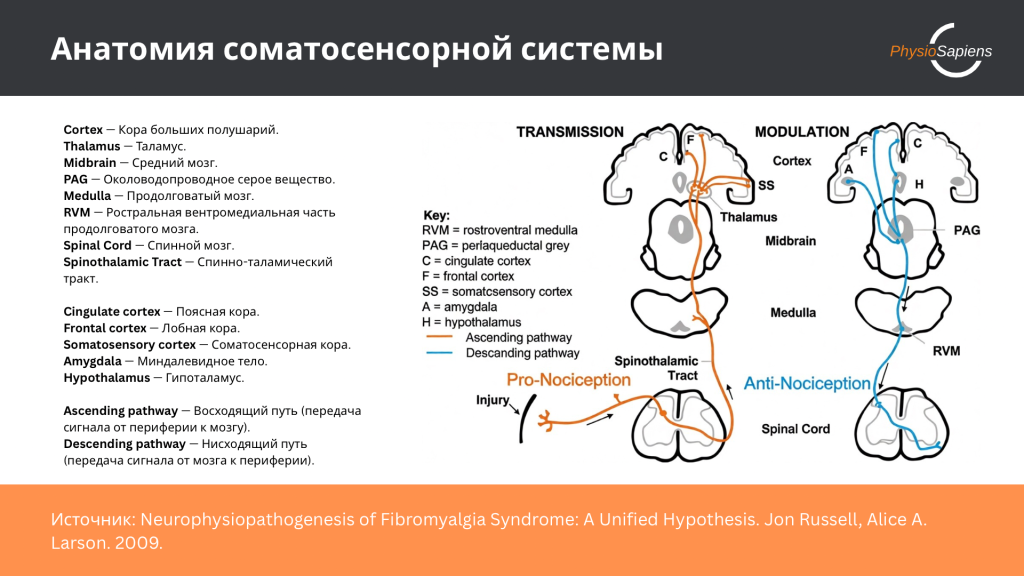
Нейрофизиологический домен боли охватывает процессы центральной обработки, модуляции и интеграции ноцицептивной информации в центральной нервной системе. Этот домен включает механизмы центральной сенситизации, нейропластичности, активацию специфических мозговых структур, функционирование нисходящих путей контроля боли и роль нейротрансмиттерных систем в модуляции болевого восприятия.
Центральная сенситизация
Центральная сенситизация представляет собой состояние повышенной возбудимости ноцицептивных нейронов в центральной нервной системе, приводящее к усилению болевых ответов на нормальные или подпороговые стимулы [11], [27]. Этот феномен играет ключевую роль в развитии хронической боли и объясняет, почему пациенты с идентичными патанатомическими изменениями могут испытывать различную интенсивность боли.
Центральная сенситизация характеризуется несколькими ключевыми проявлениями:
- гипералгезией (повышенной болевой чувствительностью к ноцицептивным стимулам);
- аллодинией (болевым ответом на неноцицептивные стимулы);
- расширением рецептивных полей ноцицептивных нейронов [11].
Опросник центральной сенситизации (CSI-R).
Исследования демонстрируют значительные индивидуальные различия в степени центральной сенситизации, что может объяснять вариабельность болевого опыта при схожих периферических патологиях [27], [28].
Нейровизуализационные исследования выявили структурные и функциональные корреляты центральной сенситизации. В частности, было показано, что индивиды с высокой степенью центральной сенситизации демонстрируют уменьшение объема хвостатого ядра и различия в активации мозговых структур в ответ на ноцицептивную стимуляцию по сравнению с индивидами с низкой сенситизацией [28]. Эти находки указывают на то, что центральная сенситизация связана с устойчивыми нейропластическими изменениями в центральной нервной системе [11], [27].
Нейропластичность и модуляция боли
Нейропластичность — это способность нервной системы к структурным и функциональным изменениям в ответ на опыт. Она играет фундаментальную роль в формировании индивидуальных различий в болевом восприятии. Хроническая боль ассоциирована с выраженными нейропластическими изменениями, включающими реорганизацию соматосенсорной коры, изменения в функционировании лимбической системы и модификацию нисходящих путей контроля боли [7], [12].
Исследования с использованием функциональной МРТ демонстрируют, что индивидуальные различия в болевой чувствительности коррелируют с различиями в активации специфических мозговых структур, включая переднюю поясную кору, инсулу, таламус, соматосенсорную кору и префронтальную кору [7], [12], [22]. Более того, вариабельность нейронной активности надежно кодирует межиндивидуальные различия в интенсивности болевого восприятия [22].
Нисходящие пути модуляции боли, включающие периакведуктальное серое вещество, ростральную вентромедиальную часть продолговатого мозга и норадренергические пути из голубого пятна, играют критическую роль в эндогенном контроле боли [26]. Индивидуальные различия в функционировании этих систем могут объяснять вариабельность в болевой чувствительности и эффективности эндогенной анальгезии [26]. Исследования показывают, что дефицит эндогенной модуляции боли является фактором риска развития хронической боли [26].
Нейротрансмиттерные системы, включая опиоидную, серотонинергическую, норадренергическую и дофаминергическую, вовлечены в модуляцию болевого восприятия [29], [30]. Вариации в генах, кодирующих компоненты этих систем, влияют на индивидуальную болевую чувствительность и ответ на анальгетическую терапию [29], [30].
Мы рекомендуем


Генетические детерминанты болевой чувствительности
Генетические факторы вносят существенный вклад в межиндивидуальную вариабельность болевого восприятия. Исследования близнецов и семейные исследования демонстрируют, что наследуемость болевых фенотипов варьирует от 30% до 60% [3], [15], [24]. Молекулярно-генетические исследования идентифицировали множество генов-кандидатов, влияющих на различные аспекты ноцицепции и болевого восприятия.
Гены ионных каналов
Гены, кодирующие ионные каналы, играют критическую роль в генерации и проведении ноцицептивных сигналов. Особое внимание исследователей привлекает ген SCN9A, кодирующий натриевый канал Nav1.7, который экспрессируется в ноцицептивных нейронах периферической нервной системы [17]. Мутации в гене SCN9A ассоциированы с широким спектром болевых фенотипов — от полной нечувствительности к боли до тяжелых хронических болевых синдромов [17].
Полиморфизмы в гене SCN9A, такие как rs6746030, связаны с индивидуальными различиями в болевой чувствительности в общей популяции [13]. Исследования демонстрируют, что носители определенных аллелей этого полиморфизма демонстрируют различную болевую чувствительность в экспериментальных условиях [13]. Другие гены ионных каналов, включая гены калиевых и кальциевых каналов, также вовлечены в модуляцию ноцицепции [17].
Гены нейротрансмиттерных систем
Ген COMT (катехол-О-метилтрансфераза), кодирующий фермент, метаболизирующий катехоламиновые нейротрансмиттеры (дофамин, норадреналин, адреналин), является одним из наиболее изученных генов-кандидатов в генетике боли [29], [30]. Функциональный полиморфизм Val158Met в гене COMT влияет на активность фермента и ассоциирован с индивидуальными различиями в болевой чувствительности, риском развития хронической боли и ответом на лечение [29], [30].
Исследования демонстрируют, что эффекты полиморфизмов COMT являются модальность-специфичными и пол-специфичными [30]. В частности, гаплотипы COMT, кодирующие низкую активность фермента, ассоциированы с повышенной чувствительностью к термической боли и капсаицин-индуцированной боли у женщин, но не у мужчин [30]. Эти находки подчеркивают сложность генетических влияний на болевое восприятие и необходимость учета взаимодействий генотип-пол и генотип-модальность боли.
Ген OPRM1, кодирующий μ-опиоидный рецептор, также играет важную роль в индивидуальных различиях болевого восприятия и ответа на опиоидную анальгезию [19]. Полиморфизм A118G в гене OPRM1 ассоциирован с различиями в болевой чувствительности и потребности в опиоидных анальгетиках, причем эти эффекты также демонстрируют пол-специфичность [19].
Полигенная природа болевой чувствительности
Современные исследования подчеркивают полигенную природу болевой чувствительности, при которой множество генов с малыми эффектами вносят вклад в формирование болевого фенотипа [15], [24], [25]. Генетическая архитектура боли включает не только отдельные гены-кандидаты, но и сложные взаимодействия между генами (эпистаз), а также взаимодействия генов с факторами окружающей среды [15], [24].
Исследования генома в целом идентифицировали множество локусов, ассоциированных с различными болевыми фенотипами [4], [15]. Эти исследования подтверждают, что генетическая предрасположенность к боли является результатом кумулятивного эффекта множества генетических вариантов, каждый из которых вносит небольшой вклад в общую вариабельность [15], [24].
Важно отметить, что генетические эффекты на болевое восприятие не являются детерминистическими, а скорее модулируют индивидуальную восприимчивость к боли во взаимодействии с факторами окружающей среды и жизненным опытом [9], [24]. Эта концепция взаимодействия генов и среды является ключевой для понимания индивидуальных различий в болевом восприятии [9].
Жизненный опыт и психосоциальные факторы
Наряду с генетическими факторами, жизненный опыт индивида и психосоциальные переменные оказывают существенное влияние на формирование болевого восприятия. Биопсихосоциальная модель боли постулирует, что болевой опыт формируется через взаимодействие биологических, психологических и социокультурных факторов [1], [2].
Стресс и эмоциональные факторы
Стресс и эмоциональное состояние играют важную роль в модуляции болевого восприятия. Хронический стресс может приводить к изменениям в функционировании гипоталамо-гипофизарно-надпочечниковой оси и симпатической нервной системы, что влияет на болевую чувствительность и способствует развитию центральной сенситизации [2], [9]. Исследования демонстрируют, что индивиды с высоким уровнем стресса имеют повышенную болевую чувствительность и больший риск развития хронической боли [2].
Эмоциональные факторы, включая тревогу, депрессию и негативный аффект, тесно связаны с болевым восприятием [2], [12]. Нейровизуализационные исследования показывают, что эмоциональная обработка боли вовлекает структуры лимбической системы, включая переднюю поясную кору и инсулу, которые также участвуют в обработке эмоций [12]. Это анатомическое и функциональное перекрытие объясняет тесную связь между эмоциональным состоянием и болевым опытом.
Важно отметить, что взаимодействие между генетическими факторами и стрессом играет критическую роль в формировании индивидуальных различий в болевом восприятии [2], [9]. Например, генетические вариации в генах стресс-реактивности могут модулировать влияние стрессовых событий на развитие хронической боли [9].
Катастрофизация боли и когнитивные процессы
Катастрофизация боли — это когнитивный процесс, характеризующийся преувеличением угрозы боли, руминацией о болевых ощущениях и чувством беспомощности в отношении боли. Она является одним из наиболее значимых психологических факторов, влияющих на болевой опыт [2]. Высокий уровень катастрофизации боли ассоциирован с повышенной интенсивностью боли, большей инвалидизацией и худшими результатами лечения [2].
Исследования демонстрируют, что катастрофизация боли связана с усилением активации мозговых структур, вовлеченных в обработку боли, включая переднюю поясную кору, префронтальную кору и инсулу [12]. Более того, катастрофизация боли может взаимодействовать с генетическими факторами в формировании болевого восприятия [2]. Например, было показано, что эффекты определенных генетических полиморфизмов на болевую чувствительность модулируются уровнем катастрофизации боли [2].
Другие когнитивные процессы, включая внимание к боли, ожидания относительно боли и убеждения о боли, также влияют на болевое восприятие [2], [12]. Эти когнитивные факторы могут модулировать активность нисходящих путей контроля боли, влияя на интенсивность болевого опыта [12].
Инструмент скрининга «желтых флагов» OSPRO — версия из 17 пунктов.
Социокультурные влияния
Социокультурные факторы, включая этническую принадлежность, культурные нормы и социальный контекст, оказывают влияние на болевое восприятие и болевое поведение [1], [2]. Исследования демонстрируют этнические различия в болевой чувствительности, распространенности хронической боли и ответе на лечение [1], [16]. Эти различия могут отражать как генетические вариации между этническими группами, так и культурные различия в отношении к боли и болевому поведению [16].
Социальная поддержка и социальные отношения также влияют на болевой опыт. Исследования показывают, что социальная поддержка может оказывать анальгетический эффект, модулируя активность мозговых структур, вовлеченных в обработку боли [2]. Напротив, социальная изоляция и конфликтные социальные отношения могут усиливать болевое восприятие [2].
Предыдущий болевой опыт также играет роль в формировании текущего болевого восприятия. Повторяющаяся или длительная боль может приводить к изменениям в центральной обработке ноцицептивной информации, включая развитие центральной сенситизации и изменения в эндогенной модуляции боли [11], [27]. Эти изменения могут объяснять, почему индивиды с историей хронической боли демонстрируют повышенную чувствительность к новым болевым стимулам.
Клинические примеры
Одним из наиболее убедительных доказательств важности нейрофизиологического домена и индивидуальных факторов в формировании болевого восприятия является часто наблюдаемое несоответствие между данными визуализации и субъективным болевым опытом. Клинические исследования регулярно демонстрируют случаи, когда пациенты с выраженными патанатомическими изменениями, выявляемыми при МРТ, не испытывают боли, в то время как другие пациенты с минимальными структурными изменениями страдают от интенсивной хронической боли [12].
Исследования асимптоматичных индивидов с использованием МРТ выявляют высокую распространенность структурных аномалий, таких как грыжи межпозвонковых дисков, дегенеративные изменения суставов и повреждения мягких тканей, у людей, не испытывающих боли [12]. Эти находки указывают на то, что наличие патанатомических изменений не является достаточным условием для возникновения боли.
С другой стороны, значительная часть пациентов с хронической болью демонстрирует минимальные или отсутствующие патанатомические изменения при визуализации [11], [12]. В этих случаях боль может быть обусловлена преимущественно нейрофизиологическими механизмами, включая центральную сенситизацию, дисфункцию на уровне нисходящих путей контроля боли и нейропластические изменения в центральной нервной системе [11], [27].
Нейровизуализационные исследования, использующие функциональную МРТ и структурную МРТ, выявили корреляты индивидуальных различий в болевом восприятии на уровне мозговых структур. Исследования показывают, что индивиды с высокой болевой чувствительностью демонстрируют различия в толщине коры, объеме серого вещества и функциональной активации специфических зон мозга по сравнению с индивидами с низкой болевой чувствительностью [18], [20], [23].
В частности, исследование Hansen и коллег продемонстрировало связь между площадью вторичной гипералгезии (маркера центральной сенситизации) и объемом хвостатого ядра и других структур головного мозга, релевантных для боли [18]. Индивиды с большей площадью вторичной гипералгезии демонстрировали меньший объем хвостатого ядра, что указывает на структурные корреляты центральной сенситизации [18], [28].
Исследование Erpelding и коллег выявило корреляции между толщиной коры в специфических регионах и индивидуальной чувствительностью к боли и температуре [20]. Эти находки подтверждают, что индивидуальные различия в болевом восприятии отражаются на структурных характеристиках мозга [20].
Функциональные нейровизуализационные исследования демонстрируют, что индивиды с различными вариантами вторичной гипералгезии демонстрируют различия в активации мозговых структур в ответ на ноцицептивную стимуляцию [28]. Эти различия в паттернах мозговой активации могут объяснять, почему пациенты с идентичными периферическими патологиями испытывают различную интенсивность боли [28].
Важно отметить, что нейровизуализационные исследования также выявили, что вариабельность нейронной активности надежно кодирует межиндивидуальные различия в интенсивности болевого восприятия [22]. Это указывает на то, что индивидуальные различия в болевом опыте имеют объективные нейрофизиологические корреляты, которые могут быть измерены с помощью современных методов нейровизуализации [22].
Клинические примеры несоответствия между данными МРТ и болевым опытом подчеркивают необходимость интегративного подхода к оценке и лечению боли, который учитывает не только патанатомические изменения, но и нейрофизиологические механизмы, генетические факторы и психосоциальные переменные [11], [12]. Понимание того, что боль является результатом сложного взаимодействия между периферическими и центральными механизмами, генетической предрасположенностью и жизненным опытом, имеет критическое значение для разработки эффективных персонализированных программ лечения.
Заключение
Индивидуальные различия в восприятии боли представляют собой сложный феномен, формирующийся в результате взаимодействия биологических, генетических и психосоциальных факторов. Современные данные убедительно демонстрируют, что болевое восприятие не может быть адекватно объяснено исключительно патанатомическими изменениями, выявляемыми при визуализации. Вместо этого, боль следует рассматривать как результат сложного взаимодействия между патанатомическим и нейрофизиологическим доменами.
Патанатомический домен может инициировать ноцицептивную сигнализацию, однако интенсивность и качество болевого опыта в значительной степени определяются процессами центральной обработки этой информации. Нейрофизиологический домен играет критическую роль в формировании индивидуальных различий в болевом восприятии [11], [12], [27].
Генетические факторы вносят существенный вклад в межиндивидуальную вариабельность болевой чувствительности, объясняя от 30% до 60% вариабельности болевых фенотипов [3], [15], [24].
Жизненный опыт и психосоциальные факторы также оказывают существенное влияние на формирование болевого восприятия [1], [2]. Эти компоненты взаимодействуют с генетическими предрасположенностями и нейрофизиологическими механизмами, формируя уникальную мозаику факторов, определяющих болевой опыт каждого индивида [2], [9].
Интегративный подход к пониманию боли, учитывающий взаимодействие патанатомического и нейрофизиологического доменов, генетических детерминант и влияния жизненного опыта, открывает новые перспективы для разработки более эффективных и персонализированных стратегий профилактики и лечения хронической боли. Признание того, что боль является многомерным феноменом, формирующимся на пересечении биологии, генетики и опыта, является ключевым шагом к улучшению качества жизни миллионов людей, страдающих от хронической боли.